
Tymidinkinas - Thymidine kinase
| Tymidinkinas | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Kristallstruktur av en tetramer av tymidinkinas från U. urealyticum (där monomererna är färg cyan, grön, röd respektive magenta) i komplex med tymidin ( rymdfyllningsmodell , kol = vitt, syre = rött, kväve = blått) .
| |||||||||
| Identifierare | |||||||||
| EG -nr. | 2.7.1.21 | ||||||||
| CAS -nr. | 9002-06-6 | ||||||||
| Databaser | |||||||||
| IntEnz | IntEnz -vy | ||||||||
| BRENDA | BRENDA -inträde | ||||||||
| ExPASy | NiceZyme -vy | ||||||||
| KEGG | KEGG -post | ||||||||
| MetaCyc | Metabolisk väg | ||||||||
| PRIAM | profil | ||||||||
| PDB -strukturer | RCSB PDB PDBe PDBsum | ||||||||
| Genontologi | AmiGO / QuickGO | ||||||||
| |||||||||
| Tymidinkinas | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Identifierare | |||||||||||
| Symbol | TK | ||||||||||
| Pfam | PF00265 | ||||||||||
| Pfam -klan | CL0023 | ||||||||||
| InterPro | IPR001267 | ||||||||||
| PROSIT | PDOC00524 | ||||||||||
| |||||||||||
| Tymidinkinas 1, lösligt | |||||||
|---|---|---|---|---|---|---|---|
| Identifierare | |||||||
| Symbol | TK1 | ||||||
| NCBI -gen | 7083 | ||||||
| HGNC | 11830 | ||||||
| OMIM | 188300 | ||||||
| RefSeq | NM_003258 | ||||||
| UniProt | P04183 | ||||||
| Övrig data | |||||||
| EG -nummer | 2.7.1.21 | ||||||
| Ställe | Chr. 17 q23.2-25.3 | ||||||
| |||||||
| Tymidinkinas 2, mitokondriell | |||||||
|---|---|---|---|---|---|---|---|
| Identifierare | |||||||
| Symbol | TK2 | ||||||
| NCBI -gen | 7084 | ||||||
| HGNC | 11831 | ||||||
| OMIM | 188250 | ||||||
| RefSeq | NM_004614 | ||||||
| UniProt | O00142 | ||||||
| Övrig data | |||||||
| EG -nummer | 2.7.1.21 | ||||||
| Ställe | Chr. 16 [1] | ||||||
| |||||||
Tymidinkinas är ett enzym , ett fosfotransferas (ett kinas): 2'-deoxitymidinkinas, ATP-tymidin 5'-fosfotransferas, EC 2.7.1.21. Det finns i de flesta levande celler. Det finns i två former i däggdjursceller, TK1 och TK2. Vissa virus har också genetisk information för uttryck av virala tymidinkinaser. Tymidinkinas katalyserar reaktionen:
- Thd + ATP → TMP + ADP
där Thd är (deoxi) tymidin , ATP är adenosintrifosfat , TMP är (deoxi) tymidinmonofosfat och ADP är adenosindifosfat . Tymidinkinaser har en nyckelfunktion vid syntesen av DNA och därför i celldelning , eftersom de är en del av den unika reaktionskedjan för att introducera tymidin i DNA. Tymidin finns i kroppsvätskorna till följd av nedbrytning av DNA från mat och från döda celler. Tymidinkinas krävs för verkan av många antivirala läkemedel . Det används för att välja hybridomcellinjer vid produktion av monoklonala antikroppar . Inom klinisk kemi används den som en proliferationsmarkör vid diagnos , kontroll av behandling och uppföljning av malign sjukdom , främst hematologiska maligniteter .
Historia
Införlivandet av tymidin i DNA demonstrerades omkring 1950. Något senare visades det att denna införlivning föregicks av fosforylering , och omkring 1960 renades och karakteriserades det ansvariga enzymet.
Klassificering
Två olika klasser av tymidinkinaser har identifierats och ingår i denna superfamilj: en familj grupperar tymidinkinas från herpesvirus samt cellulära tymidylatkinaser, de andra familjegrupperna TK från olika källor som inkluderar, ryggradsdjur , bakterier , bakteriofagen T4 , poxvirus , afrikansk svinpestvirus (ASFV) och Fish lymphocystis disease virus (FLDV). Det huvudsakliga kapsidproteinet av insekts iriserande virus tillhör också denna familj. Prosite -mönstret känner bara igen den cellulära typen av tymidinkinaser.
Isozymer
Däggdjur har två isoenzymer , som är kemiskt mycket olika, TK1 och TK2. Den förstnämnda hittades först i fostervävnad, den andra befanns vara rikligare i vuxen vävnad, och ursprungligen kallades de foster och vuxen tymidinkinas. Snart visade det sig att TK1 är närvarande i cytoplasman endast i väntan på celldelning (cellcykelberoende), medan TK2 ligger i mitokondrier och är cellcykeloberoende. De två isoenzymerna har olika reaktionskinetik och hämmas av olika hämmare.
De virala tymidinkinaserna skiljer sig fullständigt från däggdjursenzymerna både strukturellt och biokemiskt och hämmas av hämmare som inte hämmar däggdjursenzymerna. Generna för de två mänskliga isoenzymerna lokaliserades i mitten av 1970-talet. Genen för TK1 klonades och sekvenserades. Motsvarande protein har en molekylvikt av cirka 25 kD. Normalt förekommer det i vävnad som en dimer med en molekylvikt på cirka 50 kD. Det kan aktiveras av ATP. Efter aktivering är en tetramer med en molekylvikt runt 100 kD. Emellertid motsvarar enzymformen i cirkulationen inte det protein som kodas av genen: huvudfraktionen av det aktiva enzymet i cirkulationen har en molekylvikt på 730 kD och är troligen bunden i ett komplex till andra proteiner. Detta komplex är mer stabilt och har en högre specifik aktivitet än någon av de lägre molekylviktsformerna.
Rekombinant TK1 kan inte aktiveras och omvandlas till en tetramer på detta sätt, vilket visar att enzymet som förekommer i celler har modifierats efter syntes.
TK1 syntetiseras av cellen under S -fasen av celldelning. Efter att celldelningen är klar nedbryts TK1 intracellulärt och passerar inte till kroppsvätskor efter normal celldelning. Det finns en återkopplingsreglering av tymidinkinas verkan i cellen: tymidintrifosfat (TTP), produkten av den ytterligare fosforyleringen av tymidin, fungerar som en hämmare för tymidinkinas. Detta tjänar till att upprätthålla en balanserad mängd TTP tillgänglig för nukleinsyrasyntes, inte övermätta systemet. 5'-Aminothymidine, en giftfri analog av tymidin, stör denna regleringsmekanism och ökar därmed cytotoxiciteten hos tymidinanaloger som används som antineoplastiska läkemedel . De reaktionskinetiken av tymidin och tymidinanaloger fosforylering är komplicerad och endast delvis kända. Den totala fosforyleringen av tymidin till tymidintrifosfat följer inte Michaelis-Menten-kinetiken , och de olika fosfaterna av tymidin och uridin stör fosforyleringen av varandra. Kinetiken för TK från olika arter skiljer sig från varandras och även de olika formerna från en given art (monomer, dimer, tetramer och serumform) har olika kinetiska egenskaper.
Gener för virusspecifika tymidinkinaser har identifierats i Herpes simplex-virus, Varicella zoster-virus och Epstein-Barr-virus.
 + ATP --->
+ ATP --->
 + ADP
+ ADP
Tymidin reagerar med ATP för att ge tymidinmonofosfat och ADP.
Fungera
Tymidinmonofosfat, produkten av reaktionen katalyserad av tymidinkinas, fosforyleras i sin tur till tymidindifosfat av enzymet tymidylatkinas och vidare till tymidintrifosfat av enzymet nukleosiddifosfatkinas . Trifosfatet ingår i en DNA -molekyl, en reaktion katalyserad av ett DNA -polymeras och en komplementär DNA -molekyl (eller en RNA -molekyl i fallet med omvänt transkriptas , ett enzym som finns i retrovirus ).
Tymidinmonofosfat produceras också av cellen i en annan reaktion genom metylering av deoxyuridinmonofosfat , en produkt av andra metaboliska vägar som inte är relaterade till tymidin, av enzymet tymidylatsyntas . Den andra vägen är tillräcklig för att tillhandahålla tymidinmonofosfat för DNA -reparation. När en cell förbereder sig för att dela sig krävs en fullständig ny uppsättning av DNA, och kravet på byggstenar, inklusive tymidintrifosfat, ökar. Celler förbereder sig för celldelning genom att göra några av de enzymer som krävs under delningen. De finns normalt inte i cellerna och nedregleras och bryts ned efteråt. Sådana enzymer kallas bärgningsenzymer. Tymidinkinas 1 är ett sådant bärgningsenzym, medan tymidinkinas 2 och tymidylatsyntas inte är cellcykelberoende.
Brist
Tymidinkinas 2 används av cellerna för syntes av mitokondriellt DNA. Mutationer i genen för TK2 leder till en myopatisk form av mitokondriellt DNA -utarmningssyndrom. En annan orsak till TK 2-brist kan vara oxidativ stressinducerad S-glutationionering och proteolytisk nedbrytning av mitokondrialt tymidinkinas 2. Inget syndrom orsakat av TK1-brist är känt, troligen eftersom en defekt TK1-gen skulle leda till fosterdöd.
Tymidinkinas under utveckling
Bildandet av tetramer efter modifiering av tymidinkinas 1 efter syntes ökar enzymaktiviteten. Det har föreslagits att detta är en mekanism för reglering av enzymaktiviteten. Bildandet av tetramerer observeras efter utvecklingsstadiet av Dictyostelium. Dess användning för finreglering av DNA -syntes föreslås ha etablerats hos varmblodiga djur efter att de förgrenat sig från ryggradsdjuren. Även utvecklingen av tymidinkinasliknande enzymer i utvecklingen har studerats.
Artfördelning
Tymidinkinas finns i djur, växter, vissa bakterier, archeaner och virus. Tymidinkinaserna från poxvirus, afrikansk svinpestvirus, Herpes simplex-virus, Varicella zoster-virus och Epstein-Barr-virus har identifierats och i varierande grad karakteriserats. Enzymformen i virus skiljer sig från den hos andra organismer. Tymidinkinas finns inte i svampar .
Ansökningar
Identifiering av delande celler
Den första indirekta användningen av tymidinkinas i biokemisk forskning var identifiering av delande celler genom införlivande av radiomärkt tymidin och efterföljande mätning av radioaktivitet eller autoradiografi för att identifiera de delande cellerna. För detta ändamål ingår tritierat tymidin i tillväxtmediet. Trots fel i tekniken används den fortfarande för att bestämma tillväxttakten för maligna celler och för att studera aktivering av lymfocyter i immunologi.
PET -skanning av aktiva tumörer
Fluortymidin är en tymidin analog . Dess upptag regleras av tymidinkinas 1, och det tas därför företrädesvis upp av snabbt förökande tumörvävnad. Fluorisotopen 18 är en positronemitter som används vid positronemissionstomografi (PET). Fluor-18 radiomärkt fluortimidin F-18 är därför användbart för PET-avbildning av aktiv tumörproliferation och kan jämföras med den mer vanliga markören fludeoxyglucose (18F) . Ett standardiserat protokoll som hjälper till att jämföra kliniska studier har föreslagits.
Urval av hybridom
Hybridom är celler som erhålls genom att smälta tumörceller (som kan dela sig oändligt) och immunglobulinproducerande lymfocyter (plasmaceller). Hybridom kan expanderas för att producera stora mängder immunglobuliner med en given unik specificitet (monoklonala antikroppar). Ett problem är att särskilja hybridomen från det stora överskottet av ofusinerade celler efter cellfusionen. Ett vanligt sätt att lösa detta är att använda tymidin kinasnegativ (TK-) tumörcellinjer för fusionen. De tymidinkinas -negativa cellerna erhålls genom att tumörcellinjen växer i närvaro av tymidinanaloger som dödar de tymidinkinas -positiva (TK+) -cellerna. De negativa cellerna kan sedan expanderas och användas för fusion med TK+ plasmaceller. Efter fusion odlas cellerna i ett medium med metotrexat eller aminopterin som hämmar enzymet dihydrofolatreduktas och blockerar därmed de novo -syntesen av tymidinmonofosfat. Ett sådant medium som vanligtvis används är HAT -medium, som innehåller hypoxantin, aminopterin och tymidin. De ofusinerade cellerna från den tymidinkinasbristande cellinjen dör eftersom de inte har någon tymidinmonofosfatkälla. Lymfocyterna dör så småningom för att de inte är "odödliga". Endast hybridomen som har "odödlighet" från sin celllinje -förfader och tymidinkinas från plasmacellen överlever. De som producerar den önskade antikroppen väljs och odlas sedan för att producera den monoklonala antikroppen. Hybridomceller kan också isoleras med användning av samma princip som beskrivs med avseende på en annan gen, HGPRT, som syntetiserar IMP som är nödvändig för GMP -nukleotidsyntes i bärgningsvägen.
Studie av kromosomstruktur
Molekylär kammning av DNA -fibrer kan användas för att övervaka strukturen hos kromosomer i den spirande jästen Saccharomyces cerevisiae. Detta ger DNA -replikationsprofiler för enskilda molekyler. Detta kräver att jäststammarna uttrycker tymidinkinas, vilket vildtypsjäst inte gör, eftersom det är svampar (se förekomst). Därför måste en gen för tymidinkinas införlivas i genomet.
Klinisk kemi
Tymidinkinas är ett bärgningsenzym som bara är närvarande i väntan på celldelning. Enzymet frigörs inte från celler som genomgår normal delning där cellerna har en speciell mekanism för att bryta ner proteinerna som inte längre behövs efter celldelningen. Hos normala patienter är mängden tymidinkinas i serum eller plasma därför mycket låg. Tumörceller frigör enzym till cirkulationen, troligen i samband med avbrott i döda eller döende tumörceller. Tymidinkinashalten i serum fungerar därför som ett mått på malign proliferation, indirekt som ett mått på tumörens aggressivitet.
Terapeutiska tillämpningar
Vissa läkemedel är specifikt riktade mot delande celler. De kan användas mot tumörer och virussjukdomar (både mot retrovirus och mot andra virus), eftersom de sjuka cellerna replikerar mycket oftare än normala celler och även mot vissa icke-maligna sjukdomar relaterade till alltför snabb cellreplikation (till exempel psoriasis). Det har föreslagits att tymidinanalogers antivirala och anticanceraktivitet åtminstone delvis uppnås genom nedreglering av mitokondriellt tymidinkinas.
Cytostatik
Det finns olika klasser av läkemedel som är riktade mot tymidinmetabolism och därmed involverar tymidinkinas som används för att kontrollera cancerrelaterad celldelning. Kedjeterminatorer är tymidinanaloger som ingår i den växande DNA -kedjan, men modifierade så att kedjan inte kan förlängas ytterligare. Som analoger av tymidin fosforyleras denna typ av läkemedel lätt till 5'-monofosfater. Monofosfatet fosforyleras vidare till motsvarande trifosfat och införlivas i den växande DNA -kedjan. Analogen har modifierats så att den inte har hydroxylgruppen i 3'-positionen som krävs för fortsatt kedjetillväxt. I zidovudin (AZT; ATC: J05AF01) har 3'-hydroxylgruppen ersatts av en azidogrupp, i stavudin (ATC: J05AF04) har den tagits bort utan ersättning. AZT används som substrat i en av metoderna för bestämning av tymidinkinas i serum. Detta innebär att AZT stör denna metod och kan vara en begränsning: AZT är en standardkomponent i HAART -terapi vid HIV -infektion. En vanlig följd av AIDS är lymfom och den viktigaste diagnostiska tillämpningen av tymidinkinasbestämning är för övervakning av lymfom.

AZT
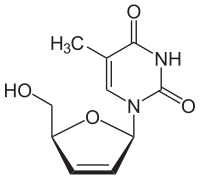
Stavudine

Idoxuridin

Aciklovir
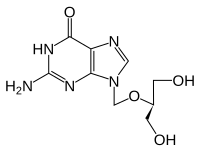
Ganciclovir





Andra tymidinanaloger , till exempel Idoxuridine (ATC: J05AB02) verkar genom att blockera basparning under efterföljande replikationscykler och gör därigenom den resulterande DNA -kedjan defekt. Detta kan också kombineras med radioaktivitet för att uppnå apoptos av maligna celler.
Antivirala medel
Vissa antivirala läkemedel, såsom acyklovir (ATC: J05AB01) och ganciklovir (ATC: J05AB06) samt andra nukleosidanaloger använder sig av substratspecificiteten för viralt tymidinkinas, i motsats till humana tymidinkinaser. Dessa läkemedel fungerar som pro-läkemedel, som i sig inte är giftiga, men omvandlas till giftiga läkemedel genom fosforylering av viralt tymidinkinas. Celler infekterade med viruset producerar därför mycket giftiga trifosfater som leder till celldöd. Humant tymidinkinas kan däremot med sin snävare specificitet inte fosforylera och aktivera förläkemedlet. På detta sätt är endast celler infekterade av viruset mottagliga för läkemedlet. Sådana läkemedel är endast effektiva mot virus från herpesgruppen med deras specifika tymidinkinas. Hos patienter som behandlas med denna typ av läkemedel observeras ofta utvecklingen av antiviralt läkemedelsresistens. Sekvensering av tymidinkinasgenen i Herpes simplex -virus och Varicella zoster -virus visar den snabba genetiska variabiliteten och kan underlätta diagnosen antiviral läkemedelsresistens.
Efter att smittkoppor förklarats utrotas av WHO i december 1979, avslutades vaccinationsprogrammen. En återuppkomst av sjukdomen antingen av en slump eller som ett resultat av biologisk krigföring möter en oskyddad befolkning och kan resultera i en epidemi som kan vara svår att kontrollera. Massvaccination för att bekämpa en koppkoppepidemi kan vara utmanande eftersom det enda godkända koppevaccinet, Vaccinia Virus, kan ha allvarliga biverkningar. Vissa regeringar lagrar dock smittkoppsvaccin för att försäkra sig om möjligheten. Utvecklingen av specifika och effektiva antivirala läkemedel prioriteras dock. Ett möjligt tillvägagångssätt skulle vara att använda specificiteten för tymidinkinas av poxvirus för ändamålet, på ett liknande sätt som det används för läkemedel mot herpesvirus. En svårighet är att poxvirus tymidinkinas tillhör samma familj av tymidinkinaser som humana tymidinkinaser och därmed är mer lik kemiskt. Strukturen av poxvirustymidinkinaser har därför fastställts för att hitta potentiella antivirala läkemedel. Sökningen har dock ännu inte resulterat i ett användbart antiviralt läkemedel mot poxvirus.
Som en "självmordsgen" i genterapi
Herpesvirustymidinkinasgenen har också använts som en "självmordsgen" som ett säkerhetssystem vid genterapiförsök, vilket gör att celler som uttrycker genen kan dödas med hjälp av ganciklovir. Detta är önskvärt om den rekombinanta genen orsakar en mutation som leder till okontrollerad celltillväxt (insertionsmutagenes). De cytotoxiska produkter som produceras av dessa modifierade celler kan spridas till närliggande celler, vilket gör dem på samma sätt mottagliga för ganciklovir, ett fenomen som kallas "åskådareffekten". Detta tillvägagångssätt har använts för att behandla cancer i djurmodeller och är fördelaktigt genom att tumören kan dödas med så få som 10% av maligna celler som uttrycker genen. Ett liknande system har prövats med användning av tomat -tymidinkinas och AZT. Dessutom används tymidinkinasgen som en självmordsgen för att ta itu med farlig transplantat-mot-värd-sjukdom vid hematopoetisk stamcellstransplantation vid namn Zalmoxis som godkändes villkorat i Europa 2016
Tumörmarkörgener
En liknande användning av tymidinkinas utnyttjar närvaron i vissa tumörceller av ämnen som inte finns i normala celler ( tumörmarkörer ). Sådana tumörmarkörer är till exempel CEA (karcinoembryoniskt antigen) och AFP (alfa -fetoprotein). Generna för dessa tumörmarkörer kan användas som promotorgener för tymidinkinas. Tymidinkinas kan sedan aktiveras i celler som uttrycker tumörmarkören men inte i normala celler, så att behandling med ganciklovir endast dödar tumörcellerna. Sådana genterapibaserade tillvägagångssätt är emellertid fortfarande experimentella, eftersom problem i samband med att rikta genöverföringen till tumörcellerna ännu inte har lösts helt.
Neutronfångsterapi för tumörer
Införlivande av en tymidinanalog med bor har föreslagits och prövats i djurmodeller för behandling av borneutroner av hjärntumörer. Ett mycket omfattande antal tymidinderivat innehållande bor har beskrivits.
Antiparasitika
Introduktion av en TK -gen i ett parasitgenom gör det möjligt att införliva BrdU och därigenom gör parasiten känslig för behandling med detta läkemedel har också föreslagits och utgör en känslig indikator på replikation av parasitgenomet.
Mått
I serum och plasma
Tymidinkinashalter i serum eller plasma har mest mätts med hjälp av enzymaktivitetsanalyser. I kommersiella analyser görs detta genom inkubering av ett serumprov med en substratanalog och mätning av mängden produkt som bildas. Direkt bestämning av tymidinkinasproteinet genom immunanalys har också använts. Mängden tymidinkinas som hittas med denna metod korrelerar inte väl med enzymaktiviteterna. En orsak till detta är att en stor mängd serum -TK1 som identifieras genom immunanalys inte är enzymatiskt aktiv. Detta är särskilt fallet med solida tumörer där immunanalyser kan vara mer känsliga.
I vävnad
Tymidinkinas har bestämts i vävnadsprover efter extraktion av vävnaden. Ingen standardmetod för extraktion eller analys har utvecklats och TK -bestämning i extrakt från celler och vävnader har inte validerats i relation till någon specifik klinisk fråga, se dock Romain et al. och Arnér et al. En metod har utvecklats för specifik bestämning av TK2 i cellextrakt med användning av substratanalogen 5-Bromovinyl 2'-deoxyuridin. I de studier som refereras nedan är metoderna som används och hur resultaten rapporteras så olika att jämförelser mellan olika studier inte är möjliga. TK1 -nivåerna i fostervävnader under utvecklingen är högre än för motsvarande vävnader senare. Vissa icke-maligna sjukdomar ger också upphov till en dramatisk höjning av TK-värdena i celler och vävnader: i perifera lymfocyter under monocytos och i benmärg under skadlig anemi. Eftersom TK1 finns i cellerna under celldelning är det rimligt att anta att TK -aktiviteten i malign vävnad bör vara högre än i motsvarande normal vävnad. Detta bekräftas också i de flesta studier.
Immunhistokemisk färgning
Antikroppar mot tymidinkinas är tillgängliga för immunhistokemisk detektion. Färgning av tymidinkinas befanns vara en tillförlitlig teknik för identifiering av patienter med stadium 2 bröstkarcinom. Det högsta antalet identifierade patienter erhölls genom kombination av tymidinkinas och Ki-67-färgning. Tekniken har också validerats för lungcancer, för kolorektalt karcinom, för lungcancer och för njurcellscancer.
Fluorescerande färgning
2'-deoxi-2 ', 2'-difluoro-5-etynyluridin (dF-EdU) binder till Herpes simplex-virus tymidinkinas men på grund av steriskt hinder, inte till humant tymidinkinas. Detta reagens tillsammans med en fluorescerande azid orsakar fluorescens av infekterade celler men inte av oinfekterade celler. Därför gör denna substratanalog det möjligt att specifikt färga infekterade celler.
Se även
- Tymidinkinas 1
- Tymidinkinas från herpesvirus
- Tymidylatkinas
- Nukleosid-difosfatkinas
- Tymidylatsyntas
- Tymidinkinas i klinisk kemi
Referenser
Vidare läsning
- Garone C, Garcia-Diaz B, Emmanuele V, Lopez LC, Tadesse S, Akman HO, et al. (Augusti 2014). "Deoxypyrimidinmonofosfat bypass -terapi för tymidinkinas 2 -brist" . EMBO Molecular Medicine . 6 (8): 1016–27. doi : 10.15252/emmm.201404092 . PMC 4154130 . PMID 24968719 .
- O'Neill KL, Buckwalter MR, Murray BK (november 2001). "Tymidinkinas: diagnostisk och prognostisk potential". Expertöversyn av Molecular Diagnostics . 1 (4): 428–33. doi : 10.1586/14737159.1.4.428 . PMID 11901857 . S2CID 12790327 .
- Topolcan O, Holubec L (februari 2008). "Tymidinkinas roll vid cancersjukdomar". Expert yttrande om medicinsk diagnostik . 2 (2): 129–41. doi : 10.1517/17530059.2.2.129 . PMID 23485133 .
- Jagarlamudi KK, Shaw M (september 2018). "Tymidinkinas 1 som en tumörbiomarkör: tekniska framsteg erbjuder en gammal biomarkör ny potential". Biomarkörer i medicin . 12 (9): 1035–1048. doi : 10.2217/bmm-2018-0157 . PMID 30039979 .
externa länkar
- Tymidin+kinas vid US National Library of Medicine Medical Subject Headings (MeSH)