
Tarmassocierad lymfoid vävnad - Gut-associated lymphoid tissue
| Tarmassocierad lymfoid vävnad | |
|---|---|
| Detaljer | |
| Systemet | Lymfsystem |
| Identifierare | |
| Akronym (er) | GALT |
| Anatomisk terminologi | |
Tarmassocierad lymfoidvävnad (GALT) är en komponent i den slemhinna-associerade lymfoida vävnaden (MALT) som fungerar i immunsystemet för att skydda kroppen från invasion i tarmen .
På grund av sin fysiologiska funktion i matabsorptionen är slemhinnans yta tunn och fungerar som en permeabel barriär mot kroppens inre. På samma sätt skapar dess bräcklighet och permeabilitet sårbarhet för infektion och i själva verket använder de allra flesta smittämnen som invaderar människokroppen denna väg. Den funktionella betydelsen av GALT i kroppens försvar är beroende av dess stora population av plasmaceller , som är antikroppsproducenter , vars antal överstiger antalet plasmaceller i mjälte , lymfkörtlar och benmärg kombinerat. GALT utgör cirka 70 viktprocent av immunförsvaret; äventyrad GALT kan påverka immunsystemets styrka som helhet avsevärt.
Strukturera
Den tarmassocierade lymfoida vävnaden ligger i hela tarmen och täcker ett område på cirka 260–300 m 2 . För att öka ytan för absorption, består tarmslemhinnan av fingerliknande utsprång ( villi ), täckt av ett monolager av epitelceller , som separerar GALT från lumentarmen och dess innehåll. Dessa epitelceller täcks av ett skikt av glykokalyx på sin luminala yta för att skydda celler från syra-pH.
Nya epitelceller som härrör från stamceller produceras ständigt på botten av tarmkörtlarna och regenererar epitelet (epitelcellens omsättningstid är mindre än en vecka). Även om konventionella enterocyter i dessa kryptor är den dominerande typen av celler, kan Paneth-celler också hittas. Dessa är placerade längst ner i kryptorna och frisätter ett antal antibakteriella substanser, bland dem lysozym, och anses vara inblandade i kontrollen av infektioner.
Under dem finns ett underliggande lager av lös bindväv som kallas lamina propria . Det finns också lymfatisk cirkulation genom vävnaden som är ansluten till de mesenteriska lymfkörtlarna .
Både GALT och mesenteriska lymfkörtlar är platser där immunsvaret startas på grund av närvaron av immunceller genom epitelcellerna och lamina propria .
GALT inkluderar också Peyers fläckar av tunntarmen , isolerade lymfoida folliklar som finns i hela tarmen och tillägget hos människor.
Följande exempel innefattar lymfoida vävnader som fungerar som gränssnitt mellan immunsystemet och inkommande antigener antingen som matantigener eller som patogena eller kommensala mikrobiotas antigener:
- Waldeyers tonsillärring
- Små lymfoida aggregat i matstrupen
- Lymfoidvävnad som ackumuleras med åldern i magen
- Peyers fläckar i tunntarmen
- Diffust fördelat lymfoida celler och plasmaceller i lamina propria i tarmen
- Intraepiteliala lymfocyter ( IEL ) isär i epitelskiktet av slemhinneytor
- Lymfoidaggregat i bilagan och tjocktarmen
- Mesenteriska lymfkörtlar som dränerar lymf kommer från tarmvävnaden
GALT kan också delas in i två kategorier med tanke på strukturen, från vilken funktionen uppstår. Det finns 1.) organiserad GALT bestående av folikler - såsom Peyers plåster, mesenteriska lymfkörtlar och ännu mer organiserad appendix. Dess huvudsakliga funktion är att inducera immunreaktion. 2.) diffus GALT med enstaka T- och B-celler, makrofager, eosinofiler, basofiler och mastceller, företrädesvis i lamina propria. Denna del av GALT består av mogna effektorceller redo att utföra sina handlingar.
Peyers plåster
Peyer-lappen är ett aggregat av lymfoida celler som projiceras mot tarmens lumen som fungerar som ett mycket viktigt ställe för initieringen av immunsvaret. Det bildar en subepitelkupol där ett stort antal B-cellsäckar med dess groddcentra, T-cellområden mellan dem i ett mindre antal och dendritceller finns. I detta område separeras subepitelkupolen från tarmlumen genom ett lager av follikelassocierat epitel . Detta innehåller konventionella tarmepitelceller och ett litet antal specialiserade epitelceller som kallas mikroveckceller (M-celler) däremellan. Till skillnad från enterocyter presenterar dessa M-celler en vikad luminal yta istället för mikrovilli, utsöndrar inte matsmältningsenzymer eller slem och saknar en tjock yta av glycocalix, så det kan vara i kontakt med mikrobiota och antigener som presenteras i tarminnehållet.
Fungera

Under normala omständigheter behöver immunförsvaret i hela organismen tarmkällor av antigener för att träna och reglera utvecklingen av olika immunceller. Utan att ha sådan stimulering utvecklas inte många egenskaper hos immunsystemet, vilket visas på fallet med bakteriefria djur. Eftersom immunceller är i konstant kontakt med bakterie- och matantigener är det primära svaret inställt som tolerogent. Fortfarande måste det finnas ett robust försvar i ett fall att patogener korsar antingen epitelgränsen eller producerar skadliga ämnen som bakterietoxiner. En sådan promenad på kanten av en kniv säkerställs av olika typer av immunceller:
B-lymfocyter
Plasma B-celler som finns vid lamina propria producerar höga nivåer av specifika sekretoriska IgA- antikroppar. Dessa IgA utsöndras i tarmens lumen genom epitelskiktet genom transcytos. För det första binder epitelceller en dimer av IgA via polymer Ig-receptor på den basolaterala sidan och transporterar den i en vesikel in i luminalutrymmet. Sedan klyvs receptorn proteolyticaly och dimeren av IgA frigörs med en del av receptorn som kallas den sekretoriska komponenten . Den sekretoriska komponenten skyddar utsöndrade antikroppar mot matsmältningsmiljön i tarmen.
En hög nivå av sekretorisk IgA härrör från interaktionen mellan B-celler och tarmantigen som presenterar dendritisk cell (DC) i samarbete med follikulär T-hjälparcell (Tfh) i germinalcentra (GC) i Peyer's lappar. Det finns två huvudsakliga sätt att producera IgA 1) T-cellberoende vilket resulterar i sIgAs med hög affinitet och specificitet och 2) T-celloberoende generation av sIgA, som använder dendritiska celler och deras produktion av BAFF- och APRIL-cytokiner. T-cellproducerade oberoende sIgAs har lägre affinitet och täcker främst kommensaler. Allmänna funktioner för sekretoriska IgA: er är att belägga någon av tarmbakterierna (kommensala eller patogena) för att försämra deras rörlighet och förhindra att de kommer i långvarig och direkt kontakt med tarmepiteliet och värdens tarmimmunsystem. Detta kallas immun utestängning. Utsöndrade IgA binder till bakterietoxiner och neutraliserar dem också.
T-lymfocyter
Naiva CD4 + T-celler differentierar till Treg eller olika hjälpar T-cellundersättningar (Th1, Th2, Th17 eller Tfh). I den tarmassocierade lymfoida vävnaden sker differentieringsprocessen via presentation av antigener härrörande från tarmmikrobiota av antigenpresenterande celler såsom dendritceller eller M-celler i Peyers plåster. Typisk process av en oral tolerans mot intagna matantigener kan beskrivas som kontinuerlig luminal provtagning av antigener av DC och deras efterföljande migration följt av priming av naiva T-celler i mesenteriska lymfkörtlar för att bli immunsuppressiva T-celler (Treg). Antigener som tas emot på detta sätt säkerställer tolerans mot dem. Kommensiell mikrobiota aktiverar också immunsvar - på ett sätt som skyddar värdens tarmvävnad från att skada den genom immuncellreaktioner. Det slutliga resultatet av T-cellpopulationer hos friska individer skiljer sig från spektrumet av T-celler till följd av akut infektion eller kronisk inflammation. Olämplig stimulering (typiskt av segmenterade filamentösa bakterier - SFB ) av kronisk Th1- eller Th17-cellrespons spelar en avgörande roll vid patologisk skada på värden.
Flera nya tarmassocierade lymfoida celler har beskrivits. De har olika ursprung och följer olika mognadsprocesser men delar samma GALT-roll - för att snabbt svara på patogener och utsöndra effektorcytokiner. Sådana flerskiktsskyddssystem belyser känsligheten hos slemhinnesäten för invaderande medel. Slemhinnan är det vanligaste sättet att införa patogena infektioner i värden. Robust ingrepp av en mängd olika lymfoida celler som patrullerar epitelskiktet återspeglar också evolutionärt tryck och kapplöpning mellan immunsystem och patogener som undgår dess kontroll.
Intraepiteliala lymfocyter (IEL)
Långlivade intraepiteliala lymfocyter är en av de största populationerna av T-lymfocyter i kroppen. Till skillnad från andra perifera lymfocyter cirkulerar inte IEL inte i blodströmmen eller lymfsystemet utan finns i epitelskiktet i tarmen. En sådan plats är mycket speciell i sig själv - IEL patrullerar tillståndet i ett enda lager av celler och har ett dendritiskt utseende. De ger den första försvarslinjen om patogen passerar epitelbarriären. IEL omfattar överraskande mångfald av ursprung och delar sig i två huvudkategorier - konventionella och icke-konventionella. Det är baserat på molekylära ytegenskaper och den plats där de får sitt slutliga cytotoxiska program för effektor.
Konventionella IEL: er har klassisk TCR-receptorer som består av α- och β-underenhet tillsammans med CD4- eller CD8-co-receptorer. De representerar en relativ minoritet av de totala intestinala IEL: erna jämfört med okonventionella IEL: er. De härrör från naiva T-celler som stöter på antigener i periferin (vanligtvis antigener som visas i DC i Peyers lappar). Efter denna aktivering uttrycker konventionella IEL-molekyler tarm-tropiska molekyler så att de kan komma hem i tarmvävnaden efter handel med lymfström och efterföljande blodström. Konventionella intraepitelceller är sålunda vävnadsbaserade effektorminnes-T-celler, som kan reagera snabbt med cytolytisk aktivitet och frisättning av cytokiner såsom INFy och TNF.
Okonventionella IEL: er presenterar majoriteten av IEL-celler i tarmepitelskiktet och förvärvar deras effektorprogram efter att ha lämnat tymus som naiva celler och stöter på antigener i GALT. Efter sådan stimulering finns okonventionella IEL (huvudsakligen γδ T-celler ) i tarmepiteliet. γδ T-celler uttrycker TCR-receptorn gjord av γ- och 5-subenhet och känner inte igen antigenpeptider som presenteras i den MHC-bundna formen. Processen för deras aktivering är fortfarande i stort sett okänd. Huvudegenskapen för γδ T-celler är bildning av långlivade minnespopulationer i barriärvävnader såsom tarmepitel eller i huden. De utför immunminnet i särskilt vävnad även efter att patogenen har rensats eller inflammationen har återtagit.
Medfödda lymfoida celler (ILC)
Innate lymfoidceller (ILC) är de mest nyligen upptäckt familj av medfödda immunceller och uttrycket ”medfödda” betyder de inte behöver antigen receptorgen ombildning. På grund av utvecklingen av en ny metod, såsom encells-RNA-sekvensering, beskrivs undertyper av medfödda lymfoida celler. Baserat på deras transkriptionsprofil inställd av huvudtranskriptionsfaktorer är ILC: er uppdelade i fem distinkta delmängder: NK-celler, ILC1, ILC2, ILC3 och LTi-celler. ILC: er förekommer vid slemhinneytor och spelar en nyckelroll i slemhinnans immunitet och homeostas på grund av deras förmåga att snabbt utsöndra immunregulatoriska cytokiner och därmed kommunicera med andra immunceller.
Medfödd immunitet
Förutom adaptiv specifik immunitet spelar en medfödd immunitet en viktig roll i GALT eftersom den ger snabbare respons. Erkännande av mikrobiell icke-självsignatur sker genom mönsterigenkänningsreceptorer (PRR) som finns på dendritiska celler, makrofager, monocyter, neutrofiler och epitelceller. PRR binder till konserverade mikrobiella mönster som cellväggskomponenter (LPS, teikoesyra, flagellin, peptidoglykan) eller viral eller bakteriell nukleinsyra. PRR är lokaliserade extracellulärt som membranbundna receptorer (Toll-liknande receptorer) eller intracellulärt (NOD-liknande och RIG-liknande receptorer). Varierad palett av mönsterigenkänningsreceptorer aktiveras av olika signaler med namnet PAMP (patogenassocierade molekylära mönster) eller av signaler kopplade till vävnadsskada med namnet DAMP (skadade associerade molekylära mönster).
Makrofager
Majoriteten av antigenpresenterande celler (APC) i tarmen härrör från makrofager, som har mycket snabb omsättning. Makrofager förekommer i lamina propria och submukosala djupare lager som muscularis-lager. Makrofager kan använda sina transepiteliala dendriter (långa cytoplasmatiska förlängningar) och direkt kontakta epitelskiktet och prova luminala bakterier. Under hälsosamma förhållanden slukar makrofager kommensbakterier och omgivande cellulärt skräp, utsöndrar IL-10, driver mognad av Treg och bidrar till vävnadshomeostas. På grund av lågt uttryck av medfödda responsreceptorer och ko-stimulerande ytmolekyler initierar inte tarmmakrofager inflammation. Men vid infektion eller inflammation förändras makrofagernas profil och de börjar utsöndra stora mängder TNF- α och bli proinflammatoriska effektorceller.
Dendritiska celler
DC-enheter presenterar mindre än 10% av lamina propria APC och finns vanligtvis inte i lägre lager. Inherent produktion av retinsyra och TGF-β (typiskt för tarmassocierade DC) inducerar expression av tarmhämmande molekyler och gynnar IgA-omkopplare under mognad av B-celler i folikuler. DCs dirigerar också Treg och konventionella IEL att ta emot sin slutliga fenotyp av mogna effektorceller i tarmen.
Andra djur
Den adaptiva immuniteten , förmedlad av antikroppar och T-celler , finns bara hos ryggradsdjur . Medan alla har en tarmassocierad lymfoidvävnad och de allra flesta har en version av mjälte och tymus , inte alla ryggradsdjur uppvisar benmärg , lymfkörtlar eller germinalcentra , vilket betyder att inte alla ryggradsdjur kan generera lymfocyter i benmärg. Denna olika fördelning av de adaptiva organen i de olika grupperna av ryggradsdjur antyder GALT som den allra första delen av det adaptiva immunsystemet hos ryggradsdjur. Det har föreslagits att från denna befintliga GALT, och på grund av trycket från kommensala bakterier i tarmen som utvecklades tillsammans med ryggradsdjur, uppträdde senare specialiseringar som tymus, mjälte eller lymfkörtlar som en del av det adaptiva immunsystemet .
Ytterligare bilder
Lymfatik i tjocktarmen.
Avsnitt av den mänskliga matstrupen.
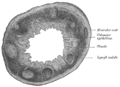
Tvärsnitt av mänsklig vermiform process. X 20.

Sektion av slemhinnan i mänskligt ändtarm. X 60.




Referenser
externa länkar
- Histologibild: 12502loa - Histology Learning System vid Boston University - "Matsmältningssystemet: matsmältningskanal: kolon , taenia coli "
- Histologibild: 11102loa - Histologins inlärningssystem vid Boston University - "Matsmältningssystemet: matsmältningskanalen: matstrupen / magkorsningen"